
Autor: Kovacs Orsi
Unha propiedade importante da biodiversidade é que non é distribuída de forma homoxénea no globo. Diferentes áreas poden albergar un número diferente de especies. Ademais, o número de especies pódese medir a diferentes escalas xeográficas: nunha fraga, nunha provincia, nun continente... E, por último, dúas áreas poden ter exactamente o mesmo número de especies, pero estas seren completamente distintas. Para definir a diversidade biolóxica a diferentes escalas, e como de diferentes son as comunidades biolóxicas na súa composición, é moi útil definir as diversidades alfa, beta e gamma.
Quen introduciu estes termos foi R.H. Whittaker en 1960. A definición máis sinxela é que a diversidade alfa é a diversidade de especies a nivel local, a diversidade gamma é diversidade de especies a nivel rexional, e a diversidade beta é a relación entre ambas. Como veremos, a diversidade beta, por tanto, reflicte o cambio na composición da comunidades. É importante recalcar que os niveis local e rexional son relativos, é dicir, podemos definir o nivel local como a fraga e o nivel rexional como a provincia, ou o nivel local como a provincia e o rexional como o continente.
A diversidade biolóxica pode medirse tamén de diferentes formas. A máis sinxela é simplemente o número de especies. Por tanto, na súa forma máis simple, definimos a diversidade alfa como o número de especies a nivel local (a media do número de especies nas diferentes localidades), a diversidade gamma como o número total de especies na rexión, e a diversidade beta como a razón entre a diversidade gama (rexional) e a diversidade alfa (local). Esencialmente, por tanto, a diversidade beta cuantifica o número de comunidades diferentes nunha rexión. Entón, está claro que a diversidade beta, ademais de ter en conta a relación entre a diversidade local ea rexional, tamén informa sobre o grao de diferenciación entre as comunidades biolóxicas. Isto é así porque as diversidades gamma e alfa serán diferentes se (e só se) as comunidades biolóxicas nunha rexión son diferentes entre localidades.
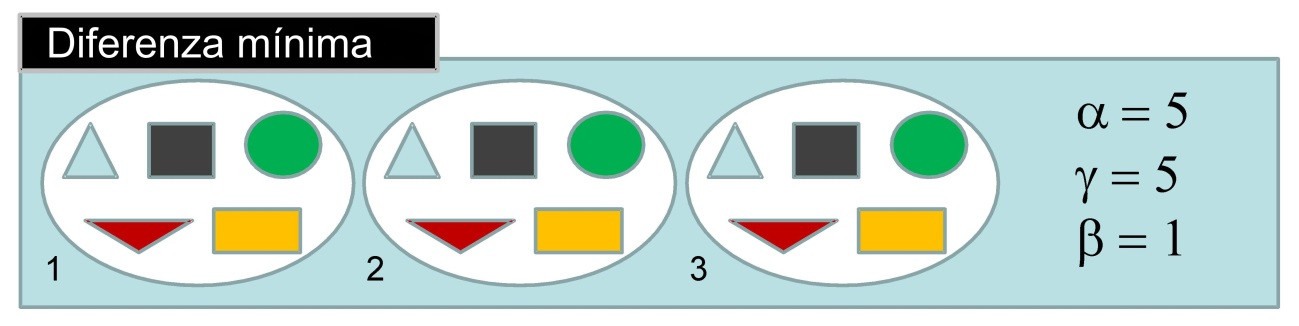
É doado demostrar como a diversidade beta varía da mínima á máxima diferenciación das comunidades locais nunha zona. Cando as comunidades locais son idénticas (diferenciación mínima), a diversidade alfa iguala a diversidade gamma e a diversidade beta é igual a 1 (Figura 1). Por tanto, beta=1 indica que neste caso, só hai unha comunidade.
Cando as comunidades locais son completamente diferentes, é dicir, presentan diferenciación máxima, a diversidade gamma é igual á multiplicación da diversidade alfa polo número de localidades (N). Entón, a diversidade beta é igual a N. Isto significa que hai N comunidades diferentes (Figura 2).
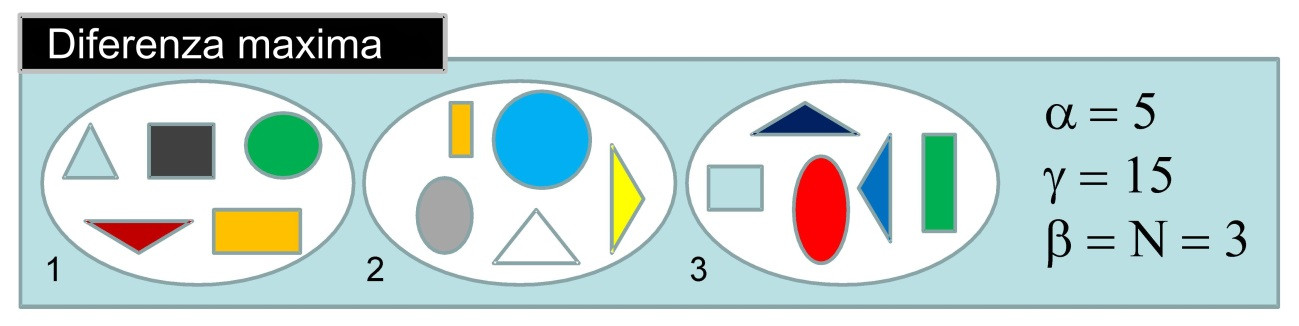
Por tanto, vemos que o valor máximo que pode tomar beta non é independente do número de localidades que hai na rexión. Entón, se queremos comparar valores de beta entre rexións que teñen diferente número de localidades, cómpre estandarizar a diversidade beta. Un exemplo é o índice de Sørensen, que é simplemente beta - 1 / N - 1. En xeral, os índices de disimilitude que son transformacións monótonas da diversidade beta (índices coma Sørensen e Jaccard, Chao et al., 2012) son medidas adecuadas de diferenzas entre as comunidades biolóxicas.
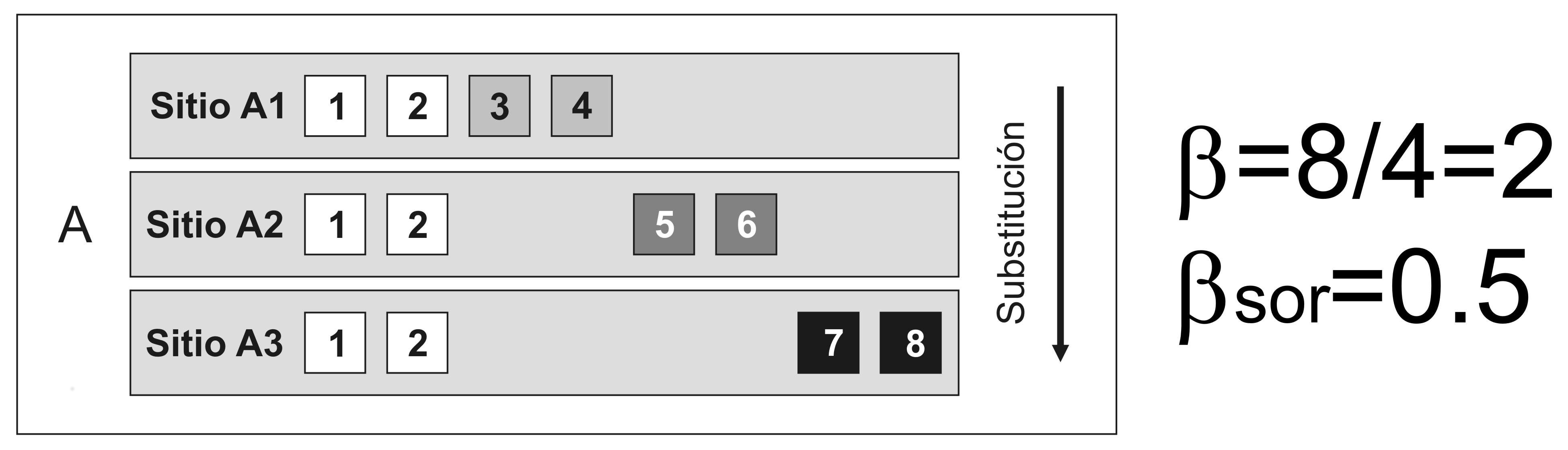
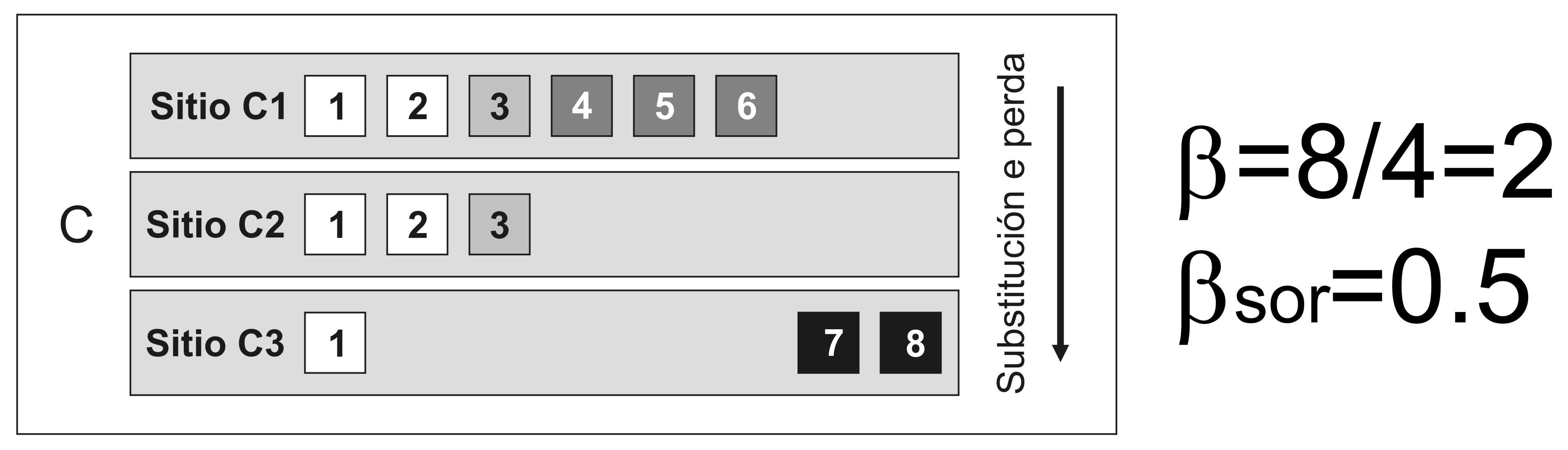
Pero, qué significa exactamente que as comunidades biolóxicas sexan diferentes? O primeiro pensamento que todos temos cando se pensa en beta diversidade ou diferenza entre comunidades biolóxicas é a substitución dalgunhas especies por outras (Figura 3).
Sen embargo, a perda de especies da comunidade máis rica á máis pobre tamén fai que as diversidades alfa e gamma sexan diferentes, potencialmente producindo o mesmo valor da diversidade beta para situacións completamente diferentes: na primeira as especies son substituídas por outras, na segunda non hai substitucións, só perda de especies (Figura 4).
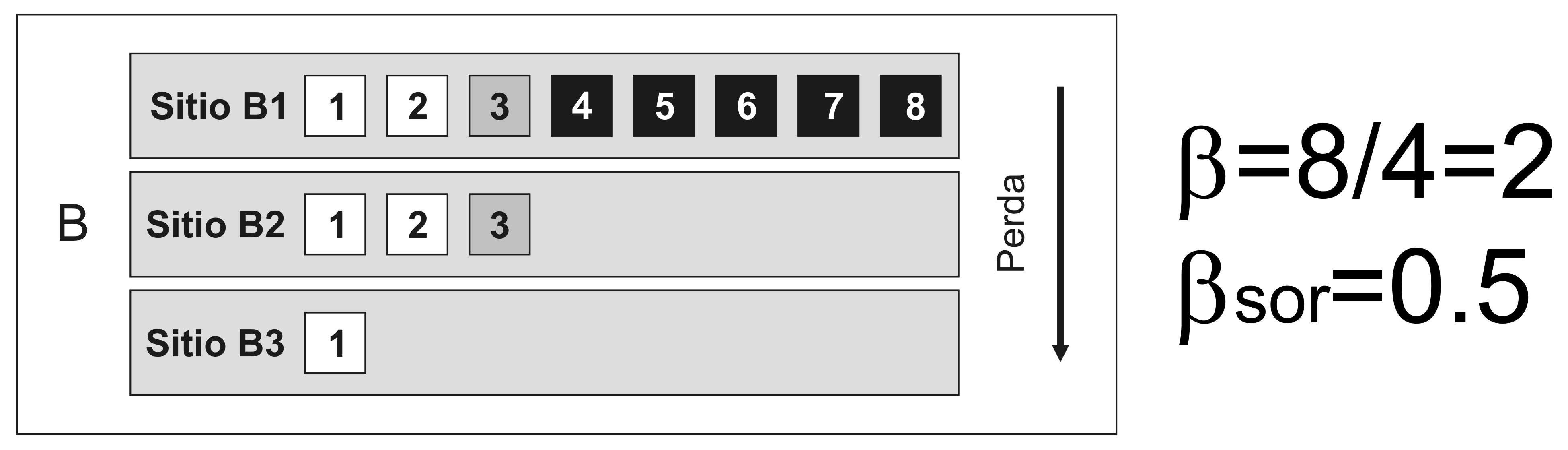 alt="">
alt="">Por suposto, nos sistemas naturais pode haber unha mestura dos dous padróns, como ilustrado na figura a continuación. Nos tres exemplos, a diversidade alfa e gama son idénticas, entón a diversidade beta e o índice de Sørensen tamén teñen os mesmos valores.
Afortunadamente, hai un xeito de separar os valores de diversidade beta producidos por estes dous padróns. O índice de disimilitude de Simpson elimina os efectos da riqueza (número de especies) sobre a diferenza entre comunidades. Este índice considera que comunidades que estean aniñadas (unha comunidade é un subconxunto doutra comunidade) son completamente similares. Por tanto, mide só os padróns de substitución de especies. Se non hai diferenza de riqueza, os índices de Simpson e Sørensen presentan o mesmo valor, o que significa que calquera diferenza entre os dous valores será debido ao compoñente aniñado da disimilitude (diferenzas de composición causadas por perda de especies). Desta forma, a disimilitude total (Sørensen) pode ser dividida aditivamente nos compoñentes de substitución e aniñamento. Podemos así relacionar as diferenzas en composición debidas a un ou outro fenómeno cos factores que os causan, respectivamente.
Todos estes métodos axudan a analizar como cambia a biodiversidade no espazo (por exemplo, por qué hai máis especies no trópico?) ou no tempo (por exemplo, diminúe a biodiversidade a medida que a paisaxe faise máis e máis humanizada?). A separación de niveis alfa, beta e gamma permite explorar cómo se producen eses cambios, a que escalas xeográficas e en relación con qué factores. Por exemplo, parece que tanto a diversidade local (alfa) e a rexional (gamma) aumentan segundo nos acercamos ao ecuador, pero a diversidade rexional faino de maneira máis acusada. Isto indica que a diversidade beta (o grao de diferenciación das comunidades) aumenta tamén segundo nos acercamos aos trópicos. Qué procesos son responsables disto? Esa é outra historia, que tratan de investigar os campos da bioxeografía e a macroecoloxía.
Escrito por Andrés Baselga
 Departamento de Zooloxía, Xenética e Antropoloxía Física. Universidade de Santiago de Compostela.
Departamento de Zooloxía, Xenética e Antropoloxía Física. Universidade de Santiago de Compostela. 


